Mitochondria
are organelles specialized for aerobic respiration. As
covered earlier, they are believed to have arisen by
endosymbiosis of a prokaryote in an ancestral eukaryotic
cell. (Mitochondria
video)
|
- The
Structure
of a Mitochondrion:
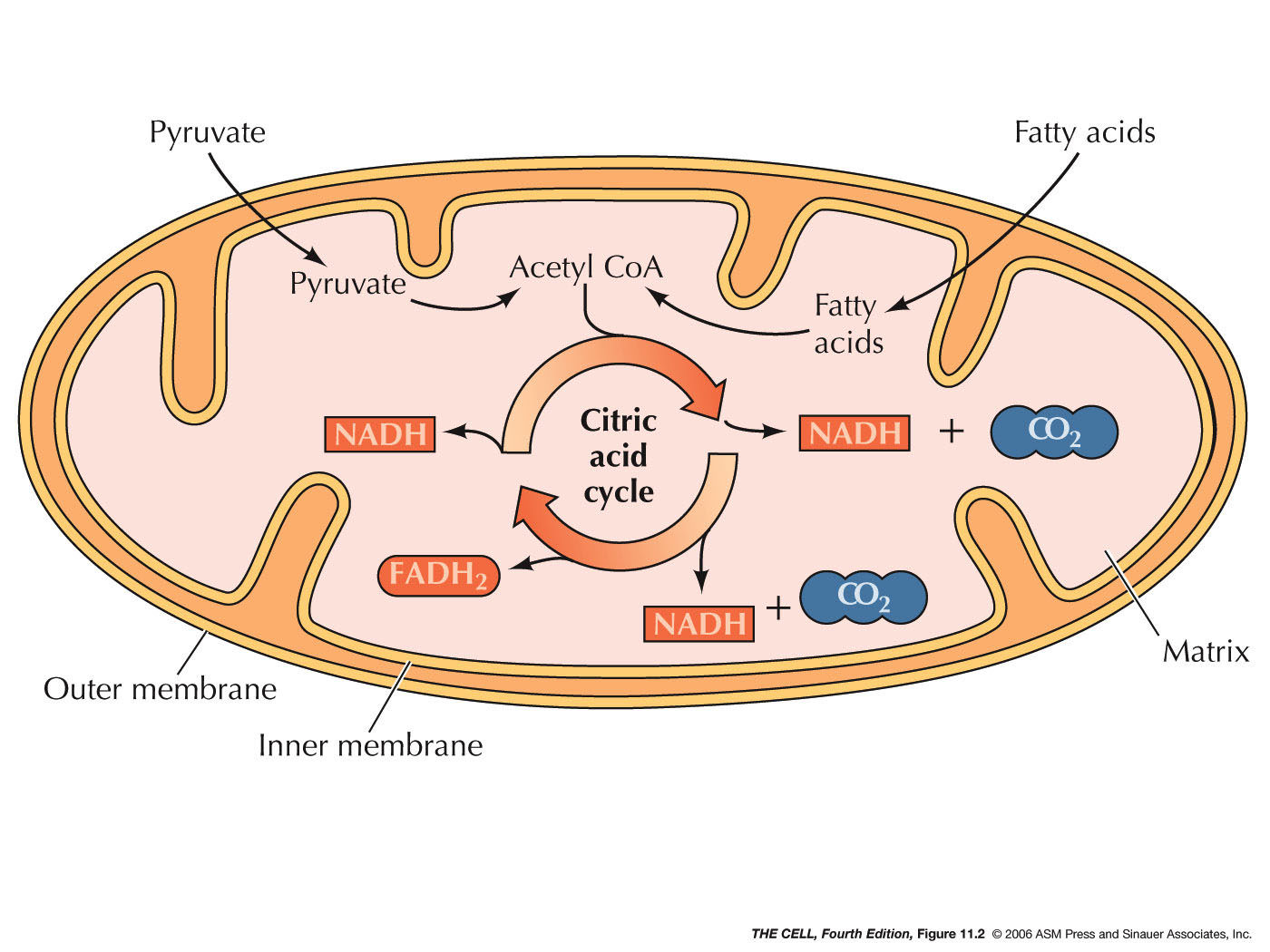
A mitochondrion has a double membrane (inner and outer)
with the inner membrane folded, creating cristae. Between
the two membranes is the intermembrane space and inside
the inner membrane is the matrix. The outer membrane is
highly permeable to small molecules due to channels
formed by porin proteins. However, the inner membrane is
impermeable to most ions and small molecules.
|

|
- The
Genetic
Material of Mitochondria: The genetic
material of mitochondria is circular DNA
(like the chromosomes of bacteria) which for most
animals is about 16 kb in length. (Plant mitochondrial
DNAs are considerable longer.) There are many copies
of this circular molecule in each mitochondrion.
Mitochondrial genes in animals include those for a few
of the proteins needed for oxidative phosphorylation
(the rest are encoded by nuclear genes)(2010 article),
mitochondrial rRNA genes, and most of the
mitochondrial tRNA genes (mt
tRNAs).
|
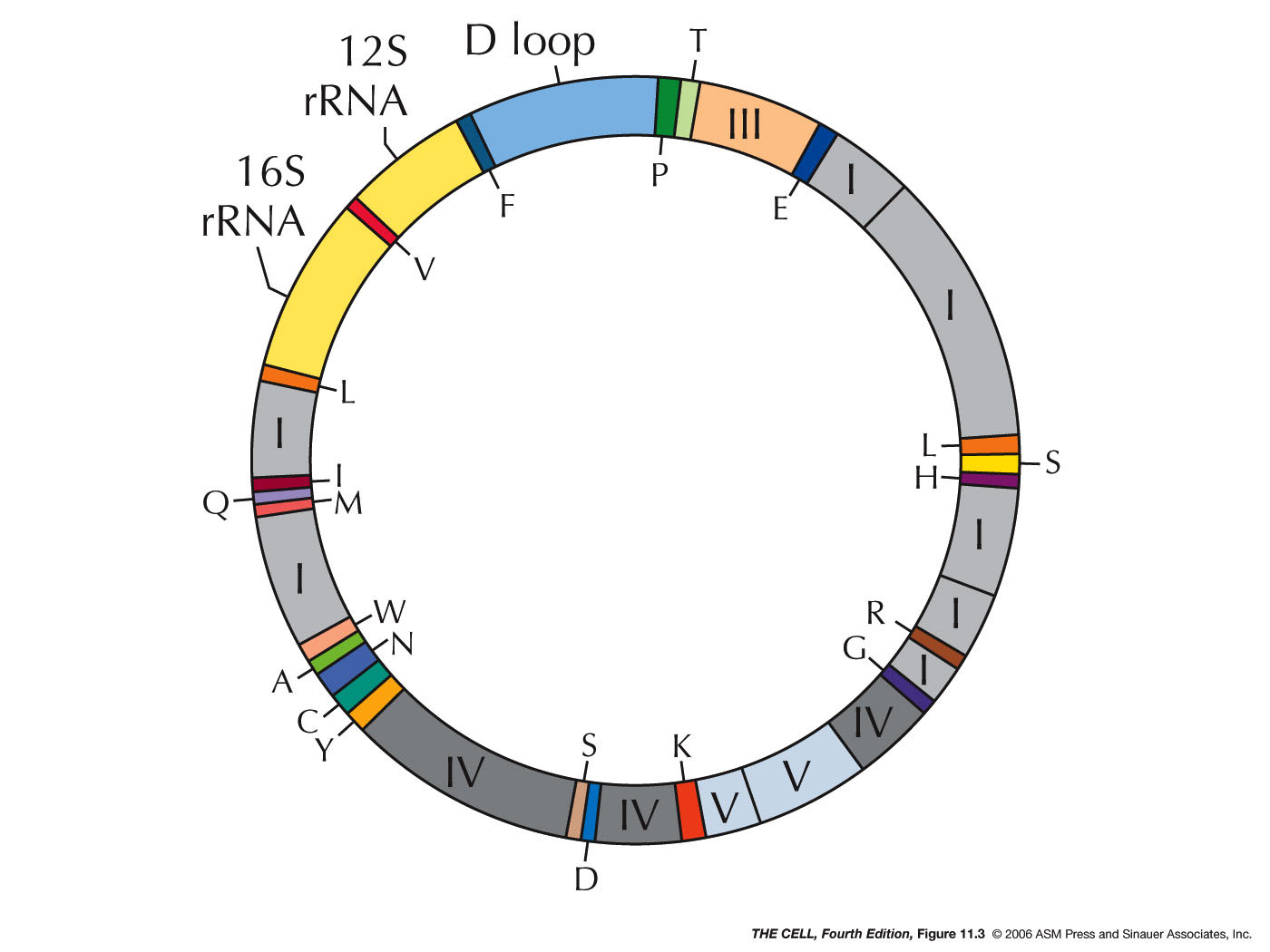 |
- The Human Mitochondrial Genome: In
humans, the mitochondrial DNA is about 16 kb and
encodes 13 proteins* that are embedded in
the inner mitochondrial membrane and are involved in
oxidative phosphorylation. It also encodes the 16S
and a 12S rRNA, and the 22 tRNAs. The 16S rRNA is
the only rRNA in the large ribosomal subunit
(39S) and the 12S rRNA is the only rRNA in the
small ribosomal subunit (28S). The mitochondrial
genetic code is and example of a deviation
from
the universal code. The ribosomal proteins
found in mitochondrial are all coded for by nuclear
genes and these proteins are imported into the
mitochodrion. The large subunit has 48 proteins, 28
of which are similar to E. coli ribosomal proteins. The
small subunit has about 30 proteins. There is a
lower RNA/protein ratio in mitochondrial versus E. coli. *But, wait a minute..."For decades,
scientists thought they had a handle on the
mitochondrial chromosome: 13 genes for proteins,
two for rRNAs and 22 for tRNAs, all tightly packed
with no introns." ... but now
"it appears to contain small open reading frames
that are hidden
inside the other genes."
- Mitochondrial Gene Inheritance:
Mitochondria are usually passed to the zygote only
via the egg and not the sperm. Therefore,
mitochondrial gene inheritance follows a unique
maternal inheritance pattern. Mitochondrial genes
also seem to have a higher mutation rate and are
therefore useful in revealing genetic differences
between closely related organisms. (See "Mutation
rate in various species" from that infallible
source Wikipedia.) Since there are multiple copies
of each mitochondrial gene in every cell, this means
that the recovery of mitochondrial genetic material
from minute samples is easier than the recovery of
nuclear genetic material.
- Endosymbiosis and Mitochondria:
Evidence for the endosymbiotic origin of
mitochondria include 1) a circular chromosome with
one origin of replication; 2) formylmethionine used
in the initiation of protein synthesis; 3)
similarity of rRNAs.
|
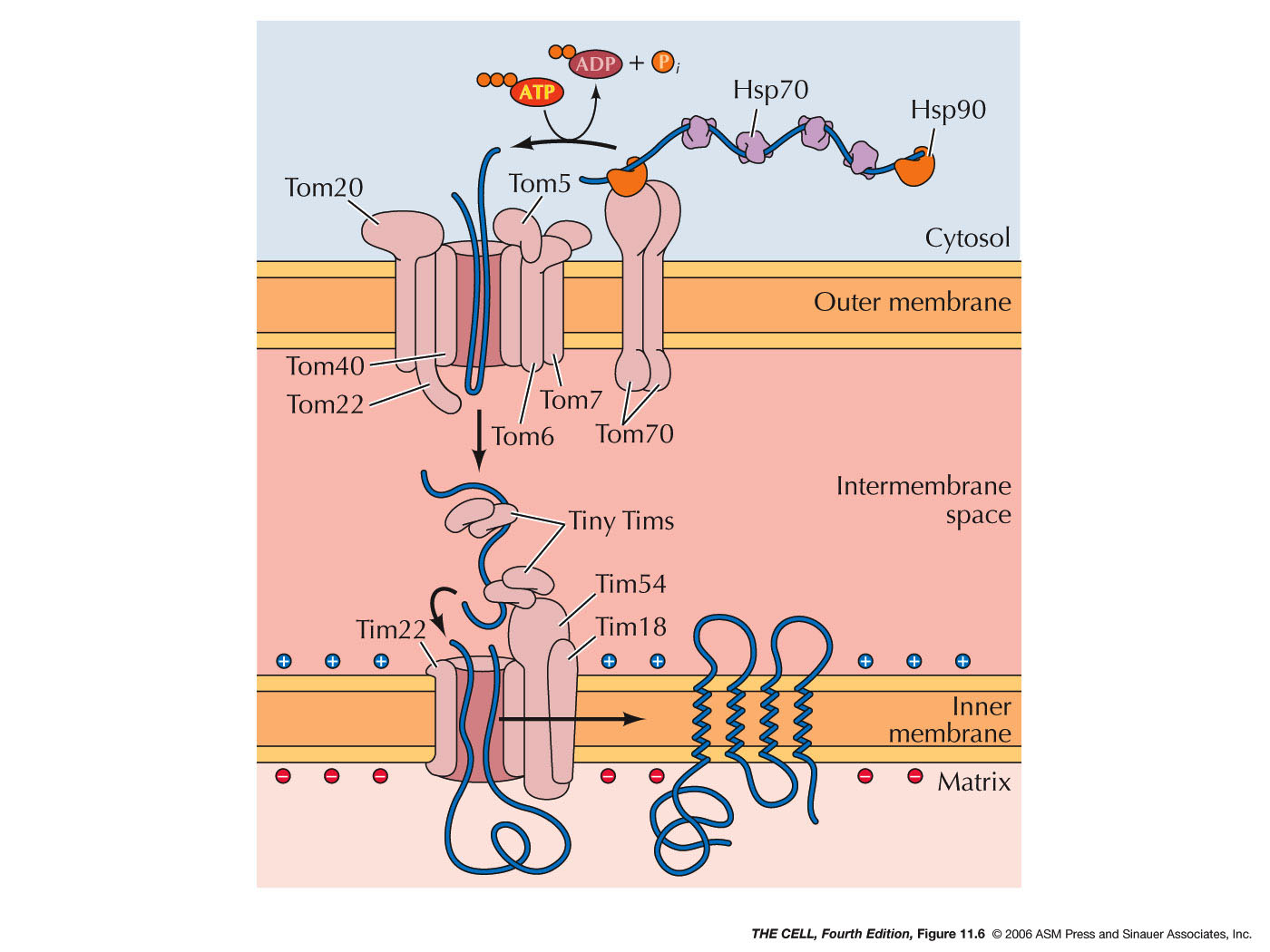
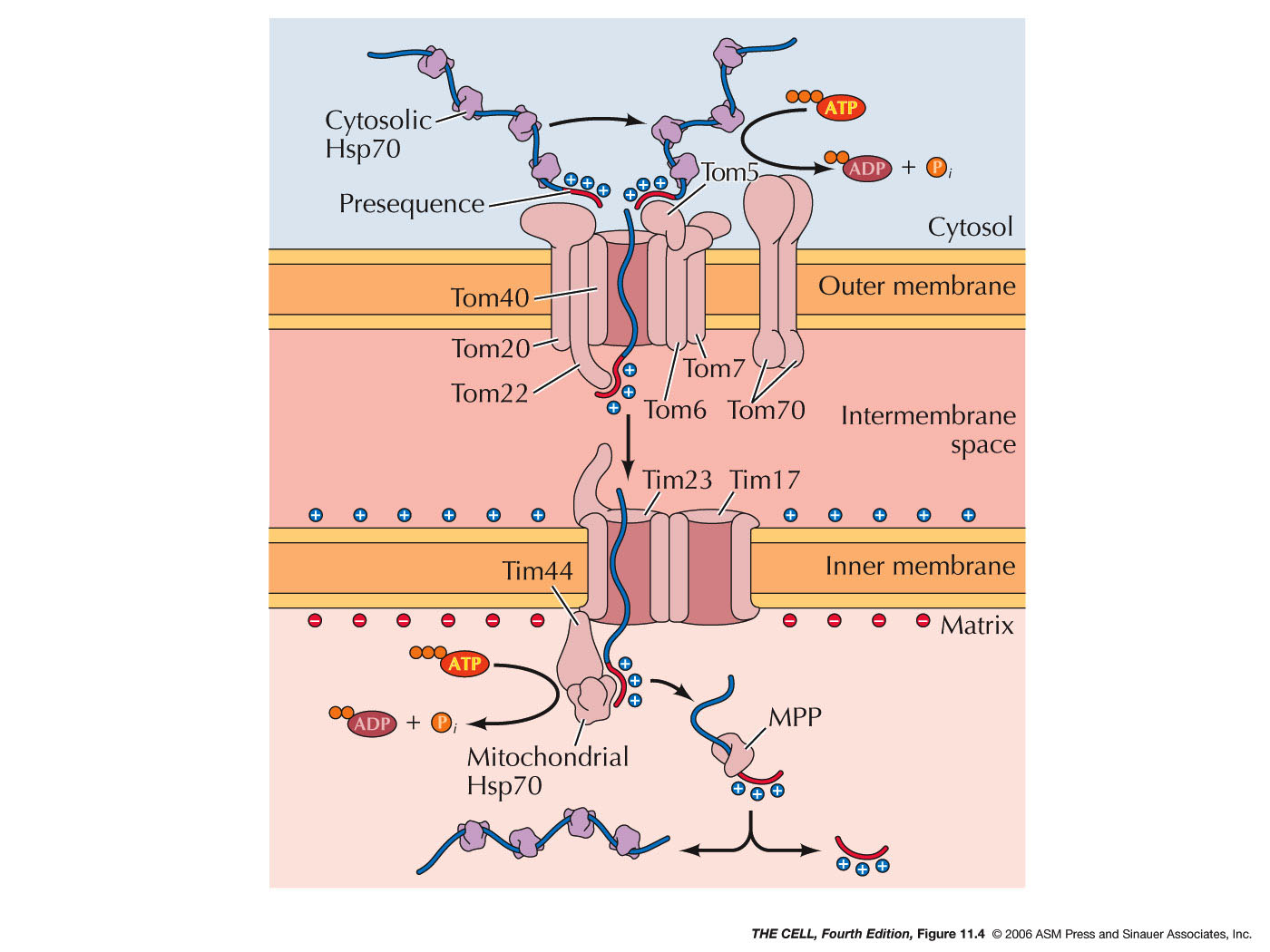
- Other
Mitochondrial Components: The vast majority
of mitochondrial proteins are imported from the
cytosol as described earlier. At least about 1000
different proteins are encoded by nuclear genes and
imported into the mitochondria. Intermembrane proteins
need only cross the outer membrane, while matrix
proteins must cross both membranes. This involves a
specific import
system similar to that required for proteins
crossing other membranes.
|
|
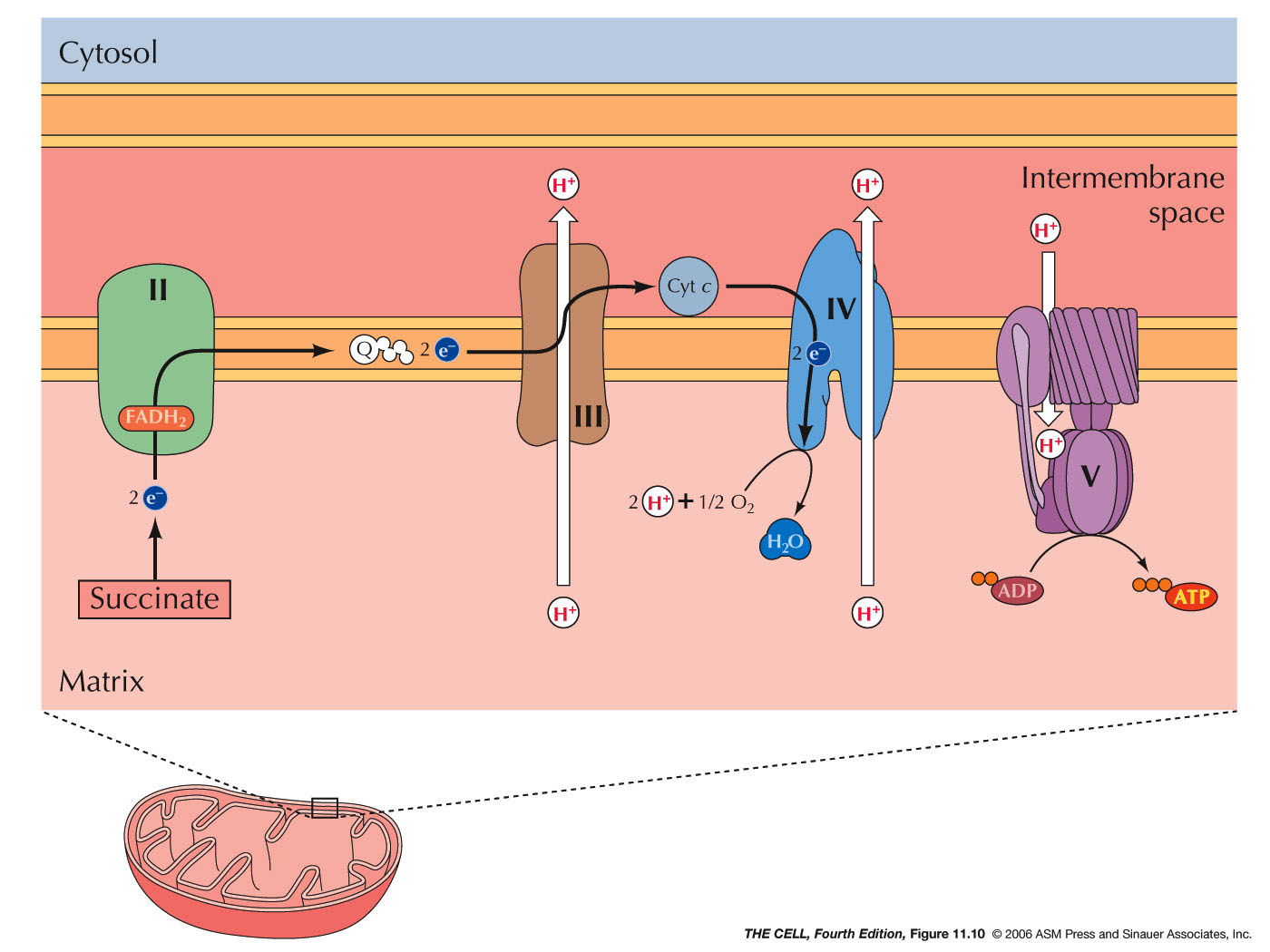
- Oxidative
Phosphorylation: During the breakdown
of energy-containing molecules, the majority of the
energy captured in the form of ATP comes from
oxidative phosphorylation. For example, the breakdown
of a molecule of glucose yields 4 ATPs directly, but
also generates 10 NADHs and 2 FADH2s
(molecules that have been reduced, receiving high
energy electrons). These electrons are then passed
through various electron transport complexes that are
embedded in the inner mitochondrial membrane. With
each transfer, energy is released and this energy is
used to pump protons (H+)
into the intermembrane space. (Some is capture
directly by the synthesis of ATP.) The proton gradient
(difference in concentration and difference in charge)
created represents potential energy and this energy is
used to manufacture ATP from ADP and P. (The
impermeability of the inner membrane maintains this
gradient.) FADH2
enters this electron transport system a little
further downstream and therefore does not produce as
great a proton gradient (fewer ATPs). The enzyme
(also in the same membrane) that catalyzed production
ATP reaction is ATP synthase. In order for this
synthesis to work, it is necessary to move ATP out of,
and ADP into the matrix. This is accomplished by a
membrane protein that exchanges one ADP in the
intermembrane space for one ATP in the matrix. The
energy to drive this transport comes from the
charge gradient (+ in intermembrane space due to the
high concentration of protons). Since ATP is more
negatively charged that ADP, this gradient favors ATP
leaving the matrix. (Electron
Transport Chain movie; ATP Synthase movie)
|
 |
 Home
Home
 Lectures
Lectures
 Videos
Videos Exams
Exams
{kind=link}
 Extra
Extra